OČI: METABOLIZAM I BIOHEMIJA VIDA
VITAMIN A/RETINOL/AKSEROFTOL
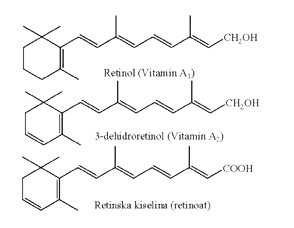
Retinol (A1) i 3-dehidroretinol (A2), dva prirodna oblika vitamina A, su C-15 izoprenoidni alkoholi sa supstituiranim β-iononskim i 3-dehidro-β-iononskim prstenom. Biološki aktivniji oblik, vitamin A1, bogato je zastupljen u jetri sisara i ribama slanih voda.
Aktivni oblici vitamina A, osim retinola su retinal (retinaldehid) i retinska kiselina. Retinol, retinal i retinska kiselina nastaju iz biljnih karotenoida (β-karoten) i pohranjuju se u jetri u obliku estara retinola sa palmitinskom, stearinskom i oleinskom kiselinom. Osim već pomenutih, izvori retinola su i žumance jajeta, maslac i neobrano mlijeko, dok se karotenoidi nalaze u tamnozelenom i žutom povrću.
Apsorpciju vitamina A i provitamina A (β-karotena) u tankom crijevu olakšavaju soli žučnih kiselina. Emulgiranje spomoću žučnih soli olakšava i hidrolizu retinil estera pomoću pankreasne retinil-ester-hidrolaze. Nakon apsorpcije slobodni retinol se u mukoznim ćelijama reesterificira sa dugolančanim masnim kiselinama, prvenstveno palmitinskom i stearinskom. Karotenoidi koji se u ćelijama cijepaju pomoću dioksigenaza, prevode se u retinal, a zadim reduciraju do retinola i esterificiraju. Retinil-esteri se zatim, asocirani sa hilomikronima, transportiraju putem limfnog sistema u jetru. Da bi ušli u parenhim, ćelije jetre, esteri retinola se ponovo hidroliziraju, vežu za celularni „retinol vezujući protein“ (RBP – retinol binding protein), reesterificiraju i pohranjuju u obliku lipoglikoproteinskih kompleksa. Da bi se oslobodio u cirkulaciju, retinol se mora odvojiti od ovog kompleksa pomoću esteraza i vezati za plazmatski „retinol vezujući protein“. U cirkulaciji, nadalje, ovaj kompleks se asocira sa prealbuminom dajući agregate (Mr ~ 55 000), koji ne mogu proći glomerularni filter.
Vitamin A ima mnogobrojne uloge u organizmu. β-karoten i drugi karotenoidi su veoma važni antioksidansi u uslovima niskog parcijalnog pritiska oksigena. Na taj način smanjuju rizik nastanka raka usljed djelovanja slobodni radikala i drugih jakih oksidanata. Retinol se u tijelu prevodi u retinil-fosfat koji je, slično dolihol-fosfatu, donor glikozilnih ostataka prilikom sinteze nekih glikoproteina i mukopolisaharida. Retinil-fosfat je bitan za sintezu glikoproteina koji su neophodni u regulaciji rasta i sekreciji sluzi. Retinol i retinska kiselina se vežu za specifične intracelularne receptore i preko ekspresije DNA utiču na sintezu proteina koji učestvuju u regulaciji rasta i diferencijacije ćelija. Znači da retinol i retinska kiselina djeluju slično steroidnim hormonima u regulaciji rasta i diferencijacije. Oblik vitamina A, δ11-cis-retinal učestvuje u procesu vida.
Noćno sljepilo jedan je od ranih simptoma nedostatka vitamina A. pošto se vitamin A pohranjuje u jetri, deficijencija se javlja samo nakon produžene neadekvatne ishrane.
ULOGA VITAMINA A U STVARANJU RODOPSINA
Vitamin A se nalazi u citoplazmi štapića i u pigmentnom sloju mrežnjače. Zbog toga je vitamin A uvijek dostupan za stvaranje novog retinala kada je to potrebno. S druge strane, kada postoji višak retinala u mrežnjači, on se pretvara nazad u vitamin A i na taj način smanjuje količinu pigmenta osjetljivog na svjetlost u mrežnjači. To prevođenje vitamina A i retinala igra važnu ulogu u dugoročnoj adaptaciji mrežnjače na različite intenzitete svjetlosti.
NOĆNO SLJEPILO
Javlja se kod ozbiljnog nedostatka vitamina A. Uzrok je što nema dovoljno vitamina A potrebnog za stvaranje odgovarajuće količine retinala i rodopsina. Stanje se naziva noćno sljepilo jer je količina svjetlosti u toku noći suviše mala da omogući odgovarajuće viđenje. Da bi noćno sljepilo nastalo, osoba mora prethodno da bude mjesecima na režimu ishrane siromašnom vitaminom A, jer se velike količine vitamina A deponuju u jetri i mogu biti dostupne očima. Noćno sljepilo se može u potpunosti izliječiti za manje od jedan sat intravenskom injekcijom vitaminom A.
NADRAŽAJ ŠTAPIĆA USLJED AKTIVACIJE RODOPSINA SVJETLOŠCU
Prvo treba reći da se mrežnjača oka sastoji od dvije vrste senzornih ćelija: štapića i čunjića. Receptorski potencijal štapića je hiperpolarizacijski, a ne depolarizacijski. Kada se štapići izlože svjetlosti, receptorski potencijal koji se stvara razlikuje se od gotovo svih drugih senzornih receptora. Nadražaj štapića uzrokoje povećanu negativnost membranskog potencijala, što je stanje hiperpolarizacije, koje označava više negativnosti sa unutrašnje membrane štapića nego što je to u mirovanju. To je potpuno suprotno od depolarizacije.
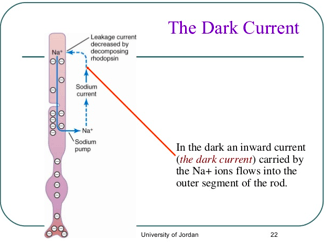
Kada se rodopsin razgradi, smanjuje se membranska provodljivost za jone natrijuma u spoljašnjem dijelu štapića. To uzrokuje hiperpolarizaciju.
Teorijska osnova za stvaranje hiperpolarizacijskog receptorskog potencijala
Unutrašnji segment neprekidno pumpa natrijum iz unutrašnjosti ka spoljašnjem dijelu štapića, stvarajući pri tome negativan potencijal u cijeloj ćeliji. Međutim, spoljašnji segment štapića u kome se nalaze diskovi fotoreceptora, je u potpunosti različit. Tu je membrana u uslovima mraka vrlo propustljiva za jone natrijuma. Zbog toga, pozitivno naelektrisani joni natrijuma neprekidno cure natrag u unutrašnjost štapića i na taj način neutrališu veliki dio negativnosti na unutrašnjoj strani čitave ćelije. Pod normalnim uslovima mraka, kada štapić nije nadražen smanjena je elektronegativnost na unutrašnjoj strani membrane štapića, oko -40mV, umjesto od -70mV do -80mV, kao u većini senzornih receptora.
Kada se rodopsin u spoljašnje segment štapića izloži svjetlosti i počne da razlaže, to smanjuje provodljivost membrane spoljašnjeg segmenta za natrijum u unutrašnjost štapića, mada se nastavlja ispumpavanje jona natrijuma kroz membranu unutrašnjeg segmenta. Više jona natrijuma napušta štapić nego što ucuri natrag u njega. To su pozitivni joni, pa njihov gubitak stvara povećanu negativnost unutar membrane, odnosno što je količina svjetlosne energije koja pogađa štapić veća i elektronegativnost postaje veća tj. veći je stepen hiperpolarizacije.
Štapići imaju hemijsku kaskadu koja amplifikuje utjecaj fotona. Pigmenti osjetljivi na boje u čepićima su kombinacija retinala i fotopsina. Nazivaju se: pigment osjetljiv na plavo, pigment osjetljiv na zeleno i pigment osjetljiv na crveno.
Talasne dužine za maksimum apsorpcije svjetlosti su: 445, 535 i 570 nm.
TRANSDUKCIJA SIGNALA U PROCESU VIDA
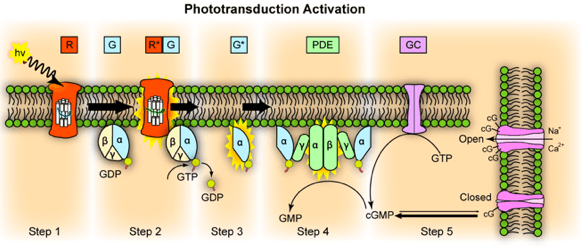
Mrežnjača oka ima dvostruku funkciju: pretvaranje svjetlosti u nervne impulse i integraciju vidne informacije. U mrežnjači su smještene fotoreceptorske ćelije šapići i čunjići. U konverziji svjetlosne energije u nervne impulse učestvuju tri međusobno povezana biohemijska ciklusa. Ovi ciklusi predstavljaju reakcije rodopsina, transducina i fosfodiesteraze, a njihov rezultat je hiperpolarizacija plazmatske membrane štapićastih ćelija. Transdukcija vida zpočinje kada svjetlost padne na rodopsin, čije su mmnogobrojne molekule smještene u membranama diskova vanjskog segmenta štapićaste ćelije. Rodopsin je protein (Mr=40 000) koji pripada familiji sedam heliksnih receptora čiji je N-terminus smješten u disku, dok je C-terminus smješten u citosolu vanjskog segmenta. Hromoforna skupina rodopsina, nalazi se u sredini dvosloja membrane diska i kovalentno je vezana za opsin, proteinsku komponentu rodopsina.
AKTIVACIJA RODOPSINA POMOĆU SVJETLOSTI
Transdukcijski proces vida sličan je prijenosu signala kod djelovanja hormona preko ćelijske membrane, a pri čemu rodopsin odgovara receptoru hormona. Nakon apsorpcije svjetla, hromoforna skupina rodopsina izomerizira u sve-trans-retinal, a rodopsin mijenja konformaciju preko serije intermedijera do metarodopsina II. Na taj način fotoreceptorska ćelija pretvara svjetlost u kretanje atoma.
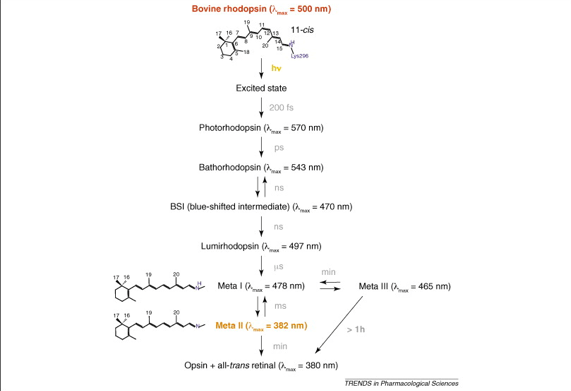
Eksitacijska „kaskada“ ukratko izgleda ovako:
1. Foton aktivira elektron u 11-cis-retinalu rodopsina, to dovodi do stvaranja metarodopsina II koji je aktivan oblik rodopsina.
2. Aktivirani rodopsin funkcionira kao enzim i aktivira veliki broj molekula transducina (protein koji se nalazi u neaktivnoj formi u membranama diskova i ćelijskoj membrani štapića).
3. Aktivirani transducin aktivira mnogo više molekula fosfodiesteraze.
4. Aktivirana fosfodiesteraza hidrolizuje mnogo molekula cGMP i tako ga razara. cGMP je prije hidrolize bio vezan za natrijumski kanal u membrani i tako ga držao otvorenim. Hidrolizom njega, na svjetlosti, natrijumski kanal se zatvara. Svaki aktiviran molekul rodopsina zatvara nekoliko stotina kanala za natrijum. Protok više od milion Na+ jona je blokiran. Smanjen tok jona Na+ ekscitira štapić.
5. Unutar sekunde,rodopsin kinaza inaktivira aktivirani rodopsin i čitava kaskada se vraća nazad na normalno stanje sa otvorenim kanalima za natrijum.
U ekscitiranoj konformaciji rodopsin (metarodopsin II) može reagirati sa proteinom transducinom koji se nalazi na citosolnoj strani membrane diska. Transducin pripada trimernoj familiji proteina koji vezuju GTP. Oni mogu vezati GTP ili GDP. U mraku se veže GDP, a sve tri podjedinice transducina (α, β i γ) su na okupu. U ovom obliku nema prenosa signala. Kada se Rh (rodopsin) pobudi pomoću svjetlosti, on reagira sa transducinom i pri tome stimulira zamjenu GDP sa GTP iz citosola. Ova zamjena dovodi do disocijacije transducina na α i βγ podjedinicu, a α, za koju je vezan GTP, prenosi signal sa ekscitiranog receptora na cGMP-fosfodiesterazu (PDE). PDE je enzim koij prevodi cGMP u 5'-GMP i specifičan je za vidne ćelije retine. PDE je integralni protein čije je aktivno mjesto na citosolnoj strani membrane diska. U mraku, čvrsto vezana inhibitorna podjedinica (I) uspješno suprimira aktivnost PDE. Kada se susretne α sa PDE, dolazi do oslobađanja inhibitorne podjedinice i povećanja aktivnosti enzima. Svaka molekula aktivirane PDE razgrađuje mnogobrojne molekule cGMP u biološki neaktivni 5'-GMP. Niža koncentracija cGMP dovodi do zatvaranja ionskih kanala ovisnih o cGMP što ima za posljedicu blokiranje pponovnog ulaska Na+ i Ca2+ u vanjski segment i hiperpolarizaciju membrane štapićastih ćelija. Na opisani način inicijalni stimulus, foton, mijenja membranski potencijal ćelije. Apsorpcija jednog fotona dovodi do zatvaranja više od 1000 ionskih kanala i mijenja membranski potencijal ćelije za oko 1mV.
Veoma brzo nakon osvjetljivanja štapićaste ćelije dolazi do građenja fotosenzornog sistema. α podjedinica T ima unutrašnju GTP-aznu aktivnost.
U vremenskom intervalu mjerenom ms nakon slabljenja intenziteta svjetlosti dolazi do hidrolize GTP i reasocijacije α sa βγ. Inhibitorna podjedinica se oslobađa i reasocira sa PDE. Na taj način dolazi do veoma jake inhibicije PDE. Da bi se koncentracija cGMP vratila na koncentraciju tamne faze, gvanilat ciklaza prevodi GTP u cGMP u reakciji koju inhibiraju visoke koncentracije Ca2+ (>100 nmol). Tokom osvjetljivanja, nivo Ca2+ pada zato što je njegova koncentracija u vanjskom segmentu rezultat ispumpavanja Ca2+ putem Na+, Ca2+ izmjene u plazmatskoj membrani i ulaska Ca2+ kroz otvorene kanale koji su ovisni o cGMP. U mraku koncentracija Ca2+ se poveća na 500 nmola što je dovoljno da se inhibira sinteza cGMP. Smanjenje nivoa Ca2+ na svjetlu dovodi do aktivacije gvanilat ciklaze i enzim prevodi GTP u cGMP što vraća sistem u stanje prije stimulusa.
Nakon produženog izlaganja svjetlosti, Rh doživljava konformacione promjene koje se ogledaju u izlaganju nekoliko Ser i Thr ostataka u C-terminalnom domenu molekule. Ovi aminokiselinski ostaci se brzo fosforilišu i uz enzim rodopsin kinazu. Niska koncentracija Ca2+ iona i rekoverin stimulišu ovu reakciju. Na fosforilisani karboksilni terminus veže se protein arestin koji sprečava daljnju reakciju između Rh i T. Ova pojava naziva se desenzitacija ili adaptacija Rh. U toku nekoliko sekundi do minute, sve-trans-retinal oblik Rh zamjenjuje se sa 11-cis-retinalom da bi nastao Rh koji je spreman za drugi krug ekscitacije. Štapićastu ćeliju mrežnice može pobuditi samo jedan foton.
Kod ljudi postoje dvije vrste receptorskih ćelija. Prema oblicima nazvani su štapići i čunjići. Čunjići djeluju pri jakom svjetlu i odgovorni su za razlikovanje boja, a štapići djeluju pri slabom svjetlu i ne učestvuju u razlikovanju boja. U retini ljudskog oka postoji oko 3 miliona čunjića i oko milijardu štapića. Ove fotoreceptorske ćelije pretvaraju svjetlost u kretanje atoma, a potom u nervni impuls. Štapići i čunjići stvaraju sinapse sa bipolarnim ćelijama, a ove bipolarne ćelije su u interakciji s drugim nervnim ćelijama u mrežnjači. Električne signale koje stvore fotoreceptori obrađuju složeni niz nervnih ćelija u mrežnjači, a potom ih vlakna optičkog nerva vode u mozak. Iz toga se može zaključiti da mrežnjača ima dvostruku ulogu: pretvaranje svjetlosti u nervne impulse i integracija vidne informacije.
Godine 1938. Selin Hecht je otkrio da ljudsku štapićastu ćeliju može pubuditi samo jedan foton. Na kojim molekulskim principima se temelji ova iznimna osjetljivost? Štapići predstavljaju izuzetno tanke tvorbe čiji promjer iznosi oko 1 µm, a dužina oko 40 µm. Glavne funkcije štapića su jasno prostorno odjeljenje. Vanjski odsječak ili segment štapića je specijaliziran za fotorecepciju. Sadrži nakupinu od otprilike hiljadu diskova, zatvorenih izravnanih vrećica debljine oko 16 nm. U tim membranskim tvorevinama su gusto raspoređene fotoreceptorske molekule. Membrane diskova odvojene su od plazmatske membrane vanjskog segmenta ćelije. Vanjski i unutrašnji segment su povezani sa jednom cilijom. Unutrašnji segment sadrži mnogo mitohondrija i ribosoma i mjesto je u kojem se sintetizira ATP i proteini. Diskovi u vanjskom segmentu imaju životni vijek samo 10 dana i zbog toga je potrebna njihova neprekidna obnova.
Unutrašnji segment nastavlja se na jezgru, a jezgra na sinaptičko tjelašce. Ovo tjelašce stvara sinapsu sa bipolarnom ćelijom i u tjelašcu su smještene sinaptičke vezikule. Fotoreceptorski protein štapića je rodopsin.
Da bi mogla podražiti fotoreceptorsku ćeliju, svjetlost se mora prvo apsorbirati. Uz to apsorpcijska skupina mora nakon apsorpcije fotona promijeniti svoju konformaciju. Fotoosjetljiva molekula koja se nalazi u štapićima je rodpsin koji je izgrađen od opsina i prostetske grupe 11-cis –retinala. Rodopsin je transmembranski protein (Mr=38 000) s N-krajem smještenim u vodenom prostoru između diskova i C-krajem na drugoj strani membrane diska u citosolu. Aminoterminalni dio rodopsina sadrži dvije oligosaharidne jedinice kovalentno vezane za asparaginske bočne ogranke. Ti šećeri vjerovatno služe za provođenje rodopsina pri premještanju unutrašnjeg odsječka u diskove. Poput drugih eukariotskih membranskih proteina i rodopsin se sintetizira u ribosonima vezanim za endoplazmatski retikulum. Novosintetizirani protein putuje tada do Goldžijevog kompleksa, a potom do citoplazmatske membrane na bazi vanjskog segmenta. To objašnjava činjenicu da šećerne jedinice rodopsina gledaju u unutrašnjost diskova, iako su u plazma-membrani okrenute u ekstracelularni prostor. Slično drugim proteinima kojima nedostaju prostetske grupe, opsin neće apsorbirati vidljivu svjetlost. Sama boja rodopsina i njegova sposobnost da apsorbuje svjetlost zavisi od prisutnosti 11-cis-retinala koji djeluje vrlo dobro kao hromofor. Molekula 11-cis-retinala omogućava rodopsinu apsorpciju unutar širokog područja u vidljivom dijelu spektra sa maksimumom apsorpcije pri 500 nm, što odgovara Sunčevoj svjetlosti. Ekstincijski koeficijent rodopsina je veoma visok oko 40 000 cm-1mol-1. Molekula 11-cis-retinala odlikuje se povoljnim hromofornim svojstvima jer je polien. Šest naizmjeničnih jednostrukih i dvostrukih veza te molekule stvaraju dugački nezasićeni elektronski niz. Molekula 11-cis- retinala vezana je za opsin preko Schiffove baze. Aldehidna skupina 11-cis-retinala vezana je na ε-amino-skupinu specifičnog lizinskog ostatka u opsinu. Spektralna svojstva rodopsina pokazuju da je Schiffova baza protonirana. Preteča 11-cis- retinala je sve-trans-retinol (vitamin A) koji sisari ne mogu sintetizirati de novo. Sve-trans-retinol se pretvara u 11-cis-retinol u dva koraka. Najprije retinol dehidrogenaza oksidira alkoholnu skupinu retinola u aldehid uz djelovanje NADP+ kao akceptora elektrona. Zatim retinal-izomeraza izomerizira dvostruku vezu između atoma C-11 i C-12 iz trans- u cis- konfiguraciju. Nedostatak vitamina A uzrokuje noćno sljepilo i, s vremenom, propadanje vanjskih odsječaka štapićastih ćelija.
SVJETLOST IZOMERIZIRA 11-CIS-RETINAL
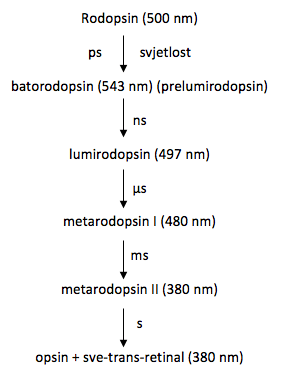
George Wald je otkrio da svjetlost izomerizira 11-cis-retinalnu skupinu rodopsina u sve-trans-retinal. Ta izomerizaciju bitno mijenja geometriju retinala. Veza preko Schiffove baze pomiče se otprilike 0,5 nm prema prstenu hromofora. Tim se događajima energija fotona pretvara u gibanje atoma. Veliki dio izomerizacije retinala se dešava unutar nekoliko pikosekundi apsorpcije fotona; to se vidi po pojavi nove apsorpcijske vrpce nakon jakog laserskog pulsa. Taj fotolitički međuprodukt, nazvan batorodopsin (ili prelumirodopsin), napet je oblik sve-trans oblika hromofora. I retinal i protein dalje mijenjaju svoje konformacije, što se vidi po stvaranju niza prolaznih međuprodukata, s posebnim spektralnim karakteristikama. Veza preko Schiffove baze deprotonira se u toku prelaska iz metarodopsina I u metarodopsin II, što traje jednu milisekundu. U toku jedne minute neprotonirana Schiffova-baza u metarodopsinu II hidrolizira se u opsin i sve-trans-retinal; opsin difuzijom napušta protein jer ne pristaje u vezivno mjesto za 11-cis-retinal. Sve-trans-retinal se u mraku izomerizira u 11-cis-izomer. Sve-trans-retinal se spaja sa opsinom i time će da regenerira rodopsin. Hidroliza Schifffove baze je prespora za razliku od reakcija fotolize rodopsina, da pridonese stvaranju živčanih impulsa. Svjetlost hiperpolarizira plazmamembranu vanjskoga odsječka.
Cis-trans izomerizacija retinala i njome izazvane konformacijske promjene rodopsina predstavljaju primarne događaje vidne pobude. Elektrofiziološka istraživanja cijelih mrežnica otkrila su sljedeći važan korak pri stvaranju živčanog impulsa: nakon svejtlosnog pulsa plazma-membrana vanjskog odsječka prolazno se hiperpolarizira. Kinetika hiperpolarizacije ovisi od intenzivnosti svjetlosnog pulsa i o razini pozadinskog osvjetljenja. Odgovor na samo jedan foton traje otprilike jednu sekundu, a jaki puls hiperpolarizira membranu za nekoliko milisekundi. Štapićasta ćelija ne stvara akcijski potencijal, već odgovara srazmjerno intenzivnosti podražaja. Svojstva signala koji su poslati iz vanjskog odsječka u sinapsu ovise o broju apsorbiranih fotona. Hiperpolarizacija potpuno osjetljivog i na tamu prilagođenog štapića dostiže polovinu najviše moguće razine, nakon što njegov vanjski odsječak koji sadrži 40X106 molekula rodopsina apsorbira samo 30 fotona. Samo jedan foton kojeg apsorbira štapić prilagođen na tamu dovodi do hiperpolarizacije od otprilike jednog mV; ta promjena se prenosi do sinapse i predaje se dalje neuronima mrežnjače. Štapići se odlikuju i nekim drugim svojstvima. Jedna od njih je ovisnost odgovora tog fotoreceptora o nivou pozadinskog osvjetljenja. Za pobuđivanje stalno osvjetljenog štapića potrebno je više fotona nego za štapić koji se nalazi u mraku. Ta se osobina naziva adaptacija i omogućava da štapićasta ćelija primijeti kontrast pri razinama pozadinskog osvjetljenja koje se razlikuje za mnogo redova veličine. Postavlja se pitanje - na kakvom se kretanju iona temelji svjetlošću izazvana hiperpolarizacija? U mraku plazma-membrana vanjskog odsječka štapića propušta Na+ ione. Zbog velikog koncentracionog gradijenta ti ioni teku brzo u vanjski odsječak. Taj gradijent održava Na+- K+- ATPazna pumpa u plazma-membrani unutrašnjeg odsječka.
Zato u mraku Na+ ioni ulaze u vanjski odsječak, difunduju u unutrašnji, odakle ih opet izbacuje pumpa koju aktivira ATP. Svjetlost nekako blokira Na+ kanale u plazma-membrani vanjskog odsječka. Zato se smanjuje ulazak Na+ iona i unutrašnjost membrane postaje negativnija. Prema tome, membranski potencijal osvijetljenog štapića pomiče se prema ravnotežnom potencijalu K+ iona. Ta svjetlost što je izazvana hiperpolarizacijom u blizini osvjetljenih diskova prenosi se tada pasivno plazma membranom do sinaptičkog tjelašca.
Međuprodukti fotolize rodopsina (označena su talasna dužina svakog apsorpcijskog maksimuma i vremenska konstanta svakog prelaska):
Signal s fotoliziranog rodopsina do plazma membarne prenose transmiteri.
Promjene propustljivosti plazma membrane na Na+ ione i time izazvana hiperpolarizacija jako su pojačani odgovori vanjskog odsječka. Apsorpcijom samo jednog fotona u štapiću adaptiranom na tamu blokira se protok više miliona Na+ iona. Membrane diskova koji sadrže većinu molekula rodopsina nisu spojene s plazma membranom štapićaste stanice. Osim toga, te membrane nisu električki spregnute. Molekula rodopsina koja apsorbira foton može biti udaljena od Na+ kanala u plazma membrani nekoliko stotina nanometara, pa je jasno da nije moguća interakcija rodopsin-kanal. Zato je gotovo sigurno da se signal s fotoliziranih rodopsina u membranama diskova do plazma-membrane prenosi difuzibilnim transmiterima. Da bi se mogao postići opaženi visok stepen pojačanja, očito je da se djelovanjem samo jednog fotoliziranog rodopsina mora stvoriti (ili razgraditi) veliki broj transmitera. Još uvijek nije otkriveno koja tvar služi kao transmiter, ali sumnja se na Ca2+ ion i ciklički GMP.
Hipotezu o Ca2+ ionu kao transmiteru potvrđuje nekoliko podataka:
* Kada nivo Ca2+ iona u citosolu poraste, Na+ kanali u plazma membrani se zatvaraju, kada se smanji, oni se otvaraju.
* Unošenjem u citosol tvari koje sa Ca2+ ionima specifično stvaraju helate snižava se osjetljivost štapićaste ćelije prema svjetlu. To smanjenje osjetljivosti upućuje na pojavu da fotoliza samo jednog rodopsina u citosolu oslobađa nekoliko stotina iona Ca2+.
* Nakon pulsa svjetlosti iz osvijetljenog vanjskog odsječka izlaze mnogi Ca2+ ioni.
Drugi eksperimenti pokazuju da ciklički GMP može biti transmiter. Glavna opažanja tome u prilog su:
* Na+ kanali u plazma membrani otvaraju se kada nivo cikličkog GMP u citosolu raste, a zatvaraju se kada njegov nivo opada.
* Nivo cikličkog GMP u ćeliji regulira svjetlo aktivacijom fosfodiesteraze koja hidrolizira taj ciklički nukleotid.
* Fotoliza samo jedne molekule rodopsina dovodi do brze hidrolize više od 105 molekula cikličkog GMP.
Svjetlost snižava koncentraciju cikličkog GMP aktivacijom fosfodiesteraze.
Eksperimenti pokazuju da i Ca2+ ioni i ciklički GMP snažno djeluju na podražljivost štapića. Sudjelovanje tih molekulskih vrsta može imati ključnu ulogu pri vidnom pobuđivanju. Još uvijek se malo zna o molekulskom mehanizmu kojim svjetlost izaziva oslobađanje Ca2+ iona u citosol. Međutim, nedavno je jako napredovalo razumijevanje kako svjetlost nadzire nivo cikličkog GMP u vanjskim odsječcima štapića. Čini se da svjetlost ne djeluje bitno na gvanil-ciklazu, enzim koji katalizira sintezu cikličkog GMP.

Nasuprot tome, svjetlost izrazito djeluje na fosfodiesterazu koja hidrolizira ciklički GMP.

Nakon osvjetljavanja, aktivnost te fosfodiesteraze poraste nekoliko stotina puta. U poticanju aktivnosti tog enzima fotoliziranim rodopsinom sudjeluje regulacijski protein transducin. U mraku transducin sadrži tijesno vezanu molekulu GDP. Pri osvjetljenju fotolizirani rodopsin stvara kompleks s GDP-transducinom i katalizira zamjenu molekule GDP molekulom GTP. Nastali GTP-transducin kompleks aktivira fosfodiesteraza. Važno je da samo jedan fotolizirani rodopsin katalizira izmjenu GDP/GTP na nekoliko stotina molekula transducina, koje opet aktiviraju stotine molekula fosfodiesteraze. Molekula GTP vezana na transducin sporo se hidrolizira i daje GDP-transducin koji više ne aktivira fosfodiesterazu.
Na taj način se slobodna energija koja daje pogon amplifikacijskom mehanizmu crpi iz hidrolize GTP. Na ovom primjeru vidi se uloga P u mehanizmu pojačanja signala.
U viđenju boja sudjeluju 3 vrste fotoreceptora.
Spektrofotometrijskim istraživanjima cijelih mrežnica otkrilo se da postoje 3 tipa čunjića koji apsorbiraju plavo, zeleno i crveno zračenje. Osvjetljivanjem čunjića zrakom svjetlosti čiji je promjer 1 mikrometar snimljeni su apsorpcijski spektri tih triju fotoreceptorskih pigmenata. Akcijski spektri za hiperpolarizaciju njihovih plazma membrana dijele se na 3 skupine. One se odlikuju apsorpcijom u plavom, zelenom, odnosno crvenom području vidljivog dijela spektra. U zlatne ribice, apsorpcijski maksimumi triju receptora za boju smješteni su pri 455, 530 i 625 nm, dok apsorpcijski maksimum rodopsina leži na 500 nm. U sve tri vrste čunjića hromofor je 11-cis-retinal. U odsustvu proteina protonirana Schiffova baza 11-cis–retinala odlikuje se apsorpcijskim maksimumom pri 380 nm. Skupine s opsina jako djeluju na hromoforska svojstva te prostetske grupe. Ovisnost apsorpcijskih svojstava 11-cis-retinala primjer je opšteg načela: interakcija s proteinom mijenja svojstva prostetske grupe. Većina oblika sljepila za boje uzrokovana je spolno vezanom recesivnom mutacijom. Otprilike 1% muškaraca slijep je na crveno, a 2% na zeleno. Spektralna mjerenja cijelog oka pokazala su da tim ljudima nedostaju molekule koje apsorbuju ili u crvenom ili u zelenom području ili pak imaju pigmente sa pomaknutim apsorpcijskim spektrom. Daltonizam, dakle, uzrokuju odsutnost ili pogreška jedne vrste opsina u čunjićima.
Očima koje razlučuju sliku odlikuju se samo organizmi svrstani u sistematske skupine mekušaca, člankonožaca i kičmenjaka. Tri vrste očiju u te tri skupine prilično se anatomski razlikuju i misli se da su se razvile neovisno. Međutim, u svojim fotoreceptorskim molekulama sve 3 sadrže 11-cis-retinal kao hromofor. To je izrazit primjer evolucijske konvergencije. Ta molekula se odlikuje jakom apsorpcijskom vrpcom, koja se može lako pomaknuti u vidljiv dio spektra. Svjetlost lako izomerizira 11-cis-retinal, dok se u mraku molekula izomerizira vrlo sporo. Izomerizacija izaziva stukturnu promjenu koja je dovoljno velika da otkoči stvaranje nervnog impulsa. Neposredne preteče 11-cis-retinala su karoteni, koji su vrlo široko rasprostranjeni u živom svijetu.
ISHRANA I VID
Nije tajna da neke namirnice dobro utiču na vid, odnosno na zdravlje očiju. Postoje mnoge stvari oko vida koje mogu krenuti naopako starenjem. Na zdravlje očiju utiču mnogi faktori kojima smo okruženi: prašina, vjetar, izduvni gasovi, fizičke traume, vibracije, dugotrajno gledanje u monitor, UV zraci i veoma niske temperature. Najopasniji faktor od njih su sunčevi ultraljubičasti zraci i to UVA i još više UVB. Naravno, dugotrajno gledanje u monitor također ima svoj negativan uticaj na zdravlje oka. Mnoge namirnice imaju blagotvoran uticaj na zdravlje očiju. Potrebne minerale, vitamine, antioksidanse i druge biološki vrijedne materije koji pogoduju boljem vidu možemo unijeti uravnoteženom ishranom bogatom lisnatim povrćem, ribom, voćem i mesom. Prvi izbor za bolji vid je uvijek mrkva. Njena snaga leži u obilju beta-karotena, likopena i luteina. Prvi je dobar za mrežnjaču oka i štiti je od UVA zračenja. Likopen djeluje kao antioksidans i štiti oči od UVB zraka. Lutein je zaštitini fitonutrijent koji ima izrazito jako antioksidativno djelovanje protiv slobodnih radikala. U tijelu se beta-karoten pretvara u vitamin A koji se opire oksidativnom oštećenju na sočivu i sprečava degenerativne promjene katarakte. Jaja su bogat izvor sumpora i cisteina, dvije komponente proteina glutationa koji čuva sočivo od oksidativnog oštećenja. Borovnice, kupine, crne ribizle i crne sorte grožđa sadrže antocijanine, koji štite i jačaju vid. To su jaki fitonutrijenti. Borovnice smanjuju zamor oka, a pored antocijanina sadrže i karotenoide, zeaksantin i lutein, kvercitin, rutin i rezveratol. Osim njih, sadrži i selen i cink. Redovna upotreba zelenog lisnatog povrća može smanjti probleme sa vidom i kataraktom. Najbolje bi bilo da se ovo povrće ne prokuhava da bi se mogli iskoristiti sve nutrijente na pravi način. Bijeli i crni luk obiluju sumpornim jedinjenjima koji potpomažu zdravlje sočiva, a sprečavaju glaukom, kataraktu i degenerativne bolesti. Osim toga, luk popravlja cirkulaciju oka i jača imunitet. Losos, tuna, sardine, bakalar i skuša sadrže omega-3-masne kiseline koje veoma povoljno utiču i na zdravlje oka. Riba je odličan izvor EPA i DHA, dvije omega-3-masne kiseline koje su važne za zdravlje ćelija oka. Bademi sadrže vitamin E koji služi za prevenciju od katarakte. Orasi su bogati omega-3-masnim kiselinama. Citrusno voće i paprike obiluju vitaminom C i odlični su u prevenciji katarakte i infekcija. Naravno, postoji i hrana koja nikako nije zdrava za oči, pa se treba potruditi izbjegavati sljedeće namirnice: šećer, trans-masti, rafinisani proizvodi, hrana sa velikim glikemijskim indeksom kao što je tjestenina, hljeb, žitarice i sve što sadrži mononatrijum glutaminat. Pretjerivanje u ovim namirnicama može biti štetno za oči i može dovesti do problema poput glaukoma.
10 ZANIMLJIVOSTI O VIDU I OČIMA
- Ako su vaše oči plave, onda dijelite gene sa svakom osobom koja ima plave oči. Prije 10 000 godina, sve oči su bile braon boje dok nije razvijena genetska mutacija koja je dovela do pojave plavih.
- Sve bebe se rađaju bez sposobnosti da razlikuju boje.
- Naše oči mogu da razlikuju preko 500 nijansi sive.
- Svako oko sadrži 107 miliona ćelija koje reaguju na svjetlost.
- Braon oči su zapravo u osnovi plave, a pigment braon boje može laserski da se ukloni i boja oka može trajno da se promijeni u plavu.
- 1 od svakih 12 muškaraca ne razlikuje boje.
- 2 posto žena ima rijetku mutaciju koja im dozvoljava da vide 100 miliona boja uz pomoć dodatne membrane na mrežnjači.
- U prosječnom životu, oko vidi 24 miliona različitih slika.
- Izraz "za treptaj oka" je nastao zato što je kapak najbrži mišić u tijelu. Treptaj traje između 100 i 150 milisekundi i moguće je trepnuti 5 puta u sekundi.
- Oko se fokusira na 50 različitih stvari u sekundi
LITERATURA
- Biokemija, Lubert Stryer
- http://www.bolestioka.com/index.php/ishrana-i-oci/prednosti-omega-3-masne-kiseline-za-oci
- http://www.kurir.rs/zabava/tech/50-neverovatnih-cinjenica-o-ljudskom-oku-zanimljivosti-koje-do-sad-niste-znali-clanak-1674968
- Medicinska fiziologija, Guyton i Hall
- http://www.ncbi.nlm.nih.gov/books/NBK10806/
- http://www.nezavisne.com/zivot-stil/zdravlje/Hrana-za-bolji-vid/234328
- http://www.optomedik.rs/ishrana-i-vid/
- Praktikum iz biohemije sa teoretskim osnovama - Lejla Begić, Selma Berbić, Zlata Mujagić, Sadik Mehikić
- http://zena.blic.rs/Zdravlje/15440/Ocuva